

You’ve all heard of carbon nanotubes. You know, really small tubes made of just carbon atoms; maybe they’re good for moving electrons around. And supposedly carbon nanotubes might play a role in the next type of computers, so-called quantum computers. That’s carbon nanotubes. Then we have Microtubules, those with biology backgrounds will know about them, but for everyone else these are also really small tubes, but inside the cells of our bodies. We’ve seen microtubules do things inside cells even using microscopes, things like giving the cell types their shape; like a scaffolding. Microtubules have also been seen helping cells divide by grabbing things like chromosomes to opposite sides of a cell, and also as rails for transporting packages around cells. Anyway, the main point is microtubules do lots of things, we know this, but the question is do they also do some of the things that carbon nanotubes do?


Looking through the microscope to see some fun things microtubules do: pull chromosomes around, give cells their shape, act as rails for transport of goods.
Not only are microtubules hollow on the inside, say around 14nm which is similar to carbon nanotubes, they are also covered in a lattice of interesting molecules that act as binary switches. This is the G-protein signaling system, and it’s not exclusive to microtubules, rather it’s a family of molecules that work together within cells to create cascades of events and state changes. The outside of microtubules have GTP/GDP, Guanosine triphosphate and diphosphate. Removal of the 3rd phosphate is one state of the switch, while GTP is the other state. This lattice of GTP/GDP on the surface of the microtubules plays a known and observed role in vesicle transport, say when a “rail cart” attaches or detaches from the tubes, as well as in cell division, as in when the tubes attach to chromosomes; but what might be most fascinating are the fun patterns they form.
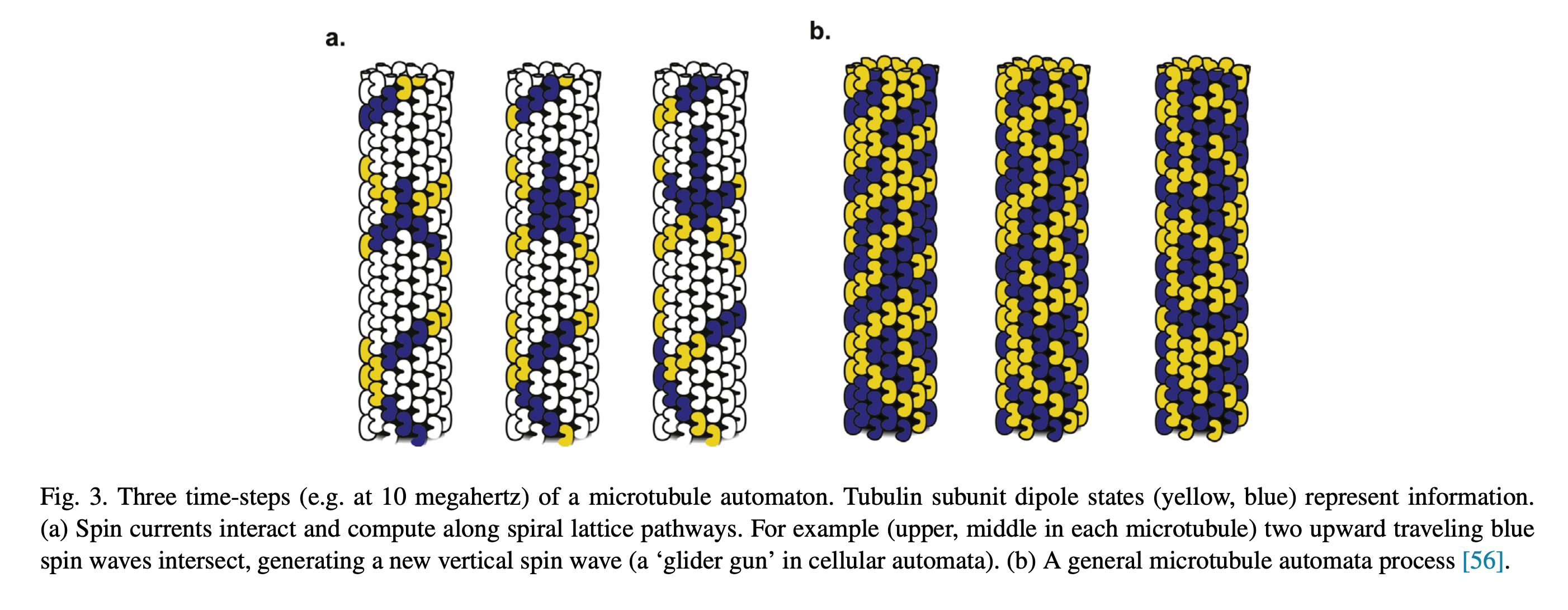
At this time for the hollow aspect of microtubules there has not been much, if any, observed behavior of electrons or other particles being transported within/through the tubes, only along the outside. But I may have just not seen those papers, or don’t know the right search terms. Some researchers have suggested, that these GTP/GDP lattices might themselves represent a form of computation, and/or electron transport. The most famous of these researchers is Roger Penrose, who is by no stretch a biologist, but there is some precedent of theoretical physicists getting close to the answer before evidence based molecular biology experiments validated the models. What comes to mind is the Trinity College lecture “What Is Life?” by Erwin Schrödinger, guessing characteristics of a molecule for transfer of hereditary information more than a decade before the structure of DNA was resolved.

Types of electron orbitals found in molecular bonds, with Pi-bonds at the bottom right, which is maybe what we are interested in.
Most bonds between atoms in molecules are either sigma bonds (single bonds) or pi-bonds (double bonds). And it seems like what Penrose and his friends are suggesting is related to a property of pi-bonds. We all know that at the center of atoms are protons & neutrons, they’re huge, and then electrons which are tiny orbit around that big center, like a planet and its moon, or a sun and its planets. But that’s really an oversimplified concept, and not really reality. As an atom gains electrons, from 1 to however many, each electron has a chance of being found in a certain region away from the center. So, yes the first 2 or 3 electrons might be found at any given time in a region that’s like concentric spheres around the atom but then things get weird. In double bonds (pi-bonds), which are common in organic molecules, a single electron can be found at the top half of the distribution or the bottom at any given instant; and when there are multiple pi-bonds in a row, like in many protein backbones, any of the electrons from any of atoms along the backbone can show up at any of the locations at any given instant. Or something, it’s weird, I don’t really get it, and maybe that’s the point Penrose is making. Quantum stuff is funky and doesn’t make much sense to dumb-dumbs like me.
There is a lot of excitement (hype) around Artificial Intelligence, all the great strides we have made with neural networks, and all the right people at the right venture retreats are saying general intelligence will change your lives forever. But much of the tech stack behind AI research is based on mimicking how neurons in biological systems are, and it may wholly be true that beyond synapses, dendrites, and neurotransmitters there is an entirely still hidden mechanism to intelligence. It may be what Penrose proposes, and it may not be. What’s clear is there is so much we do not understand about what happens within cells, on a molecular scale, and even less when it comes down to the subatomic scale. From what has been observed to date, living systems, even basic multicellular processes utilize these poorly understood mechanisms of particle behavior, let alone a process such as intelligence. Are microtubules the answer? Are carbon nanotubes and quantum processors? Is general intelligence right around the corner? Who knows. What seems important is to balance evidence based models with pure or more maths based models as guides. As is more common in physics, work it out on the chalkboard before/if ever diving in.
Further Reading:

[1] https://www.nature.com/articles/d41586-018-06166-x “The landmark lectures of physicist Erwin Schrödinger helped to change attitudes in biology”
[2] https://en.wikipedia.org/wiki/Microtubule
[3] https://en.wikipedia.org/wiki/Carbon_nanotube
[4] https://www.journals.elsevier.com/physics-of-life-reviews/news/discovery-of-quantum-vibrations



